В 2016 г. Нобелевская премия по физиологии или медицине присуждена японскому ученому Есинори Осуми (Yoshinori Ohsumi) за его открытие механизмов аутофагии. Этот процесс лежит в основе нормальной жизнедеятельности клетки. Функция аутофагии заключается в удалении частей цитоплазмы, содержащих белки, органеллы и другие составляющие. Этот процесс позволяет клетке самообновляться, а также выживать в неблагоприятных условиях. Как и все процессы на клеточном уровне, аутофагия должна пребывать в состоянии равновесия, в случае нарушения которого развиваются самые разнообразные нозологии, включая онкологические и нейродегенеративные заболевания.
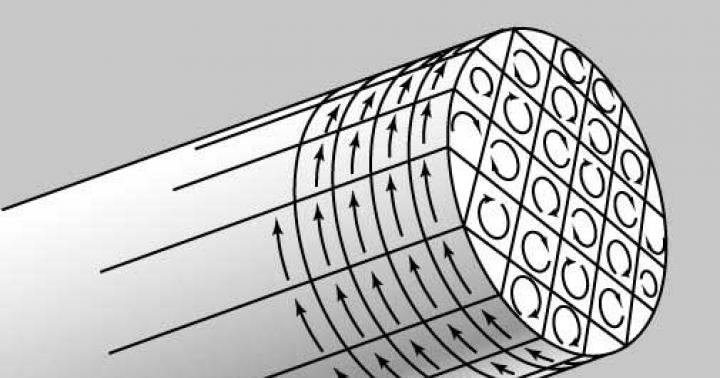
Аутофагия - эволюционно консервативный процесс, характерный для всего надцарства эукариот (организмов, содержащих ядро в клетках). Эукариотические клетки имеют возможность рециклировать части собственных составляющих путем отделения (секвестрования) части цитоплазмы двухмембранной органеллой, которая сливается с лизосомой для дальнейшего переваривания (рис. 1).
©Нобелевский комитет по физиологии или медицине (The Nobel Committee for Physiology or Medicine). Иллюстратор: Маттиас Карлен (Mattias Karlén)
В отличие от других процессов клеточной деградации, аутофагия удаляет долгоживущие белки (long-lived proteins), макромолекулярные комплексы и устаревшие или поврежденные органеллы.
Аутофагия является посредником процессов переваривания и рециклирования второстепенных (не являющихся необходимыми для выживания) частей клетки во время голодания, а также присутствует в огромном количестве физиологических процессов, в рамках которых клеточные компоненты должны быть изъяты из клетки, дабы предоставить место новым частям.
Кроме этого, аутофагия является ключевым процессом по удалению чужеродных микроорганизмов и токсических белковых агрегатов, и таким образом принимает непосредственное участие в развитии иммунного ответа и инфекционного процесса, старения и патогенеза многих заболеваний.
Несмотря на то что сам процесс аутофагии был открыт в начале 1960-х годов, механизмы и физиологическое значение оставались серым пятном клеточной биологии на протяжении нескольких десятилетий.
Что такое аутофагия
Термин «аутофагия» происходит от древнегреческого αὐτός - ауто (сам) и φαγεῖν (есть). Это процесс, при котором внутренние компоненты клетки доставляются в лизосомы или вакуоли для последующей деградации (переваривания).
В настоящее время различают три вида аутофагии.
1. Макроаутофагия. При данном процессе участок цитоплазмы, нередко содержащий органеллы, окружается мембраной, которая похожа на цистерну эндоплазматического ретикулума. Таким образом содержимое отделяется от цитоплазмы двухмембранной оболочкой. Такие двухмембранные пузырьки, внутри которых находятся органеллы и цитоплазма, называются аутофагосомами. Они соединяются с лизосомами с образованием аутофаголизосом. В них и происходит переваривание захваченного содержимого клетки. Такой процесс является как неспецифическим, так и избирательным, поскольку клетка может прибегать к этому процессу в случае необходимости избавиться от устаревших органелл (рибосом, митохондрий и прочих). Именно за открытие этого процесса присуждена Нобелевская премия в 2016 г. Далее в статье под термином «аутофагия» будет подразумеваться макроаутофагия.
2. Микроаутофагия. При этом процессе макромолекулы и частицы клеточных мембран захватываются лизосомой. Таким способом происходит переваривание белков при нехватке в клетке энергии или субстрата для синтеза новых белков. Этот процесс в целом характерен для голодания.
3. Шапероновая аутофагия. При этом процессе осуществляется целенаправленный транспорт белков (частично денатурированных) из цитоплазмы в полость лизосомы, где они подвергаются деградации. Этот тип аутофагии, насколько известно, характерен только для млекопитающих. Как правило, шапероновая аутофагия индуцируется стресс-факторами, в частности, активируется при голодании, физических нагрузках и пр. Процесс осуществляется при участии белков-шаперонов (семейства hsc-70), LAMP-2 (мембранный рецептор комплекса шаперона и транспортируемого белка) и некоторых вспомогательных белков.
Аутофагия и клеточная смерть
На сегодня ряд ученых относят аутофагию к процессам клеточной гибели. Современная классификация выделяет несколько таких процессов: апоптоз, аутофагия, некроз, аноикис (смерть клетки, вызванная ее отделением от окружающего внеклеточного матрикса), некроптоз (программируемый некроз).
С тем, что аутофагия является одним из видов клеточной смерти, согласны далеко не все. Есть множество научных исследований, подтверждающих, что аутофагия является защитным механизмом клетки, поскольку позволяет спасти ее от гибели при неблагоприятных условиях. В случае интенсификации аутофагии клетка все же погибает. Ученые выделили несколько критериев, которые позволяют говорить о том, что аутофагия является видом клеточной смерти. Так, должны отсутствовать признаки апоптоза, не должны быть активированы каспазы, конденсироваться хроматин. В клетке должно увеличиваться число аутофагосом и аутолизосом. Кроме того, ингибирование белков аутофагии должно предотвращать смерть клетки.
Процессы, связанные с клеточной гибелью, происходят постоянно и поддерживаются в строгом равновесии, сдвиг которого в одну или другую сторону неминуемо приводит к развитию нарушений и, в конечном счете, - к смерти организма. Какую роль все-таки играет в этом процессе аутофагия - ученым еще предстоит узнать.
Загадка аутофагии или предыстория открытия
Открытию Е. Осуми, за которое он получил наиболее престижную научную награду, предшествовали ранние исследования.
В 1950-е годы Кристиан Рене де Дюв (Christian Renе de Duve) изучал действие инсулина в клетках печени крыс. В рамках своей исследовательской работы он изучал локализацию фермента глюкозо-6-фосфатазы. Этот фермент играет важнейшую роль в гликогенолизе - процессе расщепления гликогена до глюкозы. В печени фермент глюкозо-6-фосфатаза расщепляет промежуточный метаболит на фосфат и глюкозу, которая затем поступает в кровь. При исследовании глюкозо-6-фосфатазы был использован метод фракционирования, разработанный Альбером Клодом (Albert Claude). Однако выявить ферментную активность в свежевыделенных печеночных фракциях не удавалось. В то же время ферментная активность сохранялась и усиливалась после того, как фракции хранились 5 сут в холодильнике.
Таким образом, Кристиан де Дюв и его студенты выявили различия в активности кислой фосфатазы в зависимости от способа выделения. Первоначально команда решила, что причина в какой-либо технической ошибке. Однако в последующих экспериментах результаты получались точно такими же.
Довольно скоро стало понятно, что ферменты каким-то образом отделены от всего содержимого клетки мембранной органеллой. На протяжении 3 лет были выявлены еще несколько кислых ферментов, связанных с этой микросомальной фракцией. Далее последовали опыты, показавшие, что фракция является самостоятельной органеллой.
В 1955 г. на основании полученных данных Кристиан де Дюв предложил название «лизосома» для органеллы, окружающей мембраной клеточные частицы, внутри которой поддерживается низкий рН (кислотный) и в которой функционируют кислые ферменты.
В том же 1955 г. Алекс Новиков (Alex B. Novikoff, американский ученый украинского происхождения) посетил лабораторию Кристиана де Дюва. Там он сделал электронные фотографии открытых органелл. Далее А. Новиков провел эксперименты и подтвердил, что в лизосомах содержится кислая фосфатаза.
За эти и другие открытия Кристиану де Дюву, Альберу Клоду и Джорджу Паладу (George E. Palade) была присуждена Нобелевская премия в 1974 г.
Интересный факт, что А. Новиков также внес большой вклад в открытие лизосом и понимание их функционирования. Однако ученый подвергался социальному и политическому преследованию. Так, в 1953 г. он был уволен из медицинского колледжа Университета штата Вермонт за отказ в сотрудничестве по раскрытию его друзей-коммунистов. Он также дважды получил отказ по поводу службы в американской армии. Спустя 20 лет после начала расследования его связей с коммунистами дело против ученого было закрыто по причине отсутствия существенных доказательств. Спустя 30 лет после начала преследований Университет принес ему официальные извинения и предоставил почетную степень доктора. В официальном заявлении Университета Миннесоты значится, что А. Новиков является одним из наиболее недооцененных ученых в контексте получения Нобелевской премии.
Вскоре после открытия лизосом ученые стали все больше узнавать об этих органеллах. Так, было показано, что определенные «порции» цитоплазмы отделяются (секвестрируются) мембранными структурами во время нормального развития почек у мышей. Выявлено, что аналогичные структуры, содержащие небольшой объем цитоплазмы и митохондрий, находятся в клетках проксимальных канальцев при гидронефрозе.
Ученые установили, что вакуоли группируются вместе с гранулами, содержащими кислые фосфатазы. Также стало известно, что количество таких структур увеличивается по мере интенсификации дегенеративных процессов.
Мембранные структуры, содержащие дегенеративную цитоплазму, находились и в нормальной печени (исследовалось на крысах), но их количество существенно возрастало при воздействии на животное глюкагоном (перфузией) или токсическими агентами.
Понимая, что эти структуры обладают свойством переваривать части внутриклеточного содержимого, Кристиан де Дюв ввел термин «аутофагия» в 1963 г. и широко описал эту концепцию в публикации, сделанной несколькими годами позже.
На протяжении следующих десятилетий достижения в этой области были весьма ограниченными. Установлено, что питательная среда и гормоны оказывают влияние на аутофагию, в частности, стимулирование инсулином супрессирует аутофагию в тканях млекопитающих.
Одно из исследований продемонстрировало, что на ранних этапах процесса происходит формирование двухмембранной структуры - фагофоры, которая образуется вокруг участка цитоплазмы и замыкается в пузырек, не содержащий гидролитических ферментов, - аутофагосому (см. рис. 1).
Открытие Осуми - поворотный этап в понимании аутофагии
Несмотря на понимание того, что аутофагия - чрезвычайно важный клеточный процесс, механизмы его функционирования и регуляции не были известны. Исследования этого процесса были довольно непростыми, поскольку период жизни фаговор и аутофагосом очень короткий. Аутофагосома существует только 10–20 мин перед слиянием с лизосомой, поэтому морфологические и биохимические исследования проводить крайне сложно. Работой в этом направлении занимались неохотно. Всего лишь несколько исследовательских лабораторий по всему миру изучали этот процесс, в основном используя сравнительные и описательные подходы и фокусируясь на поздних стадиях аутофагии, то есть непосредственно перед слиянием с лизосомой.
В начале 1990-х годов, почти через 30 лет после введения Кристианом де Дювом термина «аутофагия», еще не были выделены молекулярные маркеры и не были описаны компоненты (этапы) механизма аутофагии. Множество фундаментальных вопросов оставались открытыми: инициация процесса, формирование аутофагосом, роль аутофагии в клеточном и организменном выживании, влияние на патогенез заболеваний.
В это время Е. Осуми, тогда ассистент профессора в Токийском университете, принял решение изучать аутофагию, используя дрожжи Saccharomyces cerevisae в качестве модельной системы.
Первое, что решил узнать ученый, - существует ли аутофагия в этих одноклеточных организмах. Вакуоли дрожжей являются эквивалентом лизосом млекопитающих. Е. Осуми предположил, что если аутофагия существует у дрожжей, ингибирование вакуольных ферментов приведет к накоплению поглощенных цитоплазматических компонентов в вакуоли. Для того чтобы проверить выдвинутую гипотезу, он создал штамм дрожжей, у которых отсутствовали вакуольные протеазы: протеаза А, протеаза В и карбоксипептидаза.
Он выявил, что аутофагические тела накапливаются в вакуоле при росте дрожжей в обедненной питательной среде. Такие вакуоли становились очень большими, и их можно было исследовать под световым микроскопом.
Е. Осуми идентифицировал уникальный фенотип, который стал использовать для открытия генов, регулирующих индукцию аутофагии. Ученый индуцировал случайные мутации. И одна из них оказалась первой удачей: он смог найти первый ген, при мутации которого не отмечено накопления аутофагических телец, то есть не происходят начальные этапы процесса аутофагии. Этот ген он назвал - ген аутофагии 1 (autophagy gene 1 - APG-1 ). Продолжая работу в том направлении, ученому удалось выявить 75 мутаций.
В 1993 г. он опубликовал статью, в которой сообщил об открытии 15 генов, играющих ключевую роль в активации аутофагии в эукариотических клетках. Они получили название APG-1–15 . После открытия этих генов у других организмов всей группе было присвоено название ATG -гены. Эта номенклатура используется вплоть до настоящего времени.
На протяжении последующих лет Е. Осуми клонировал некоторые ATG -гены и охарактеризовал их функции. Клонирование гена ATG-1 позволило узнать, что он кодирует серин/треониновую киназу. Таким образом, было доказано, что в процессе аутофагии происходит фосфорилирование белков (присоединение фосфатного остатка, что является одним из самых распространенных механизмов посттрансляционной модификации белка, приводящей к изменению его активности).
Дополнительные исследования показали, что ATG-1 формирует комплекс с белком, кодируемым геном ATG-13 , и что это взаимодействие регулируется TOR-киназой (рис. 2).

TOR-киназа регулирует клеточный рост и выживание. Она активна в клетках, растущих в среде, богатой питательными веществами. Она фосфорилирует ATG-13 и препятствует формированию комплекса ATG-13:ATG-1 . Так, при голодании TOR-киназа инактивируется и дефосфорилированный ATG-13 соединяется с ATG-1 .
ATG-13:ATG-1 соединяется с ATG-17, ATG-29 и ATG-31 . Образование этого пентамерного комплекса является первым этапом в каскаде реакций, необходимых для формирования аутофагосомы.
Формирование аутофагосомы также требует наличия интегрального белка (пронизывающего мембрану) - ATG-9, фермента фосфатидилинозитола-3-киназы (PI3K) и ряда других протеинов (Vps-34, Vps-15, ATG-6 и ATG-14).
Для формирования мембраны требуются дополнительные белки и молекула фосфатидилинозитол-3 фосфата. Расширение мембраны с образованием зрелой аутофагосомы происходит при активности двух каскадов убиквитинсопряженных реакций.
В вопросах клеточного функционирования большое значение имеет локализация белков. Е. Осуми сделал интереснейшее открытие: в нормально растущих клетках дрожжей белок ATG-8 распределяется равномерно по всей цитоплазме. А в случае голодания этот же белок формирует крупные агрегаты, которые скапливаются в аутофагосомах и аутофаготических тельцах.
Ученому принадлежит еще одно удивительное открытие. Оказалось, что мембранная локализация гена ATG-8 зависит от двух увиквитинсопряженных реакций, которые работают последовательно. Они приводят к ковалентному (наиболее крепкому) присоединению ATG-8 к липидной мембране, а именно - к молекуле фосфатидилэтаноламина. Обе системы активируются одним и тем же ферментом - ATG-7.
Во время первого этапа ATG-12 активируется путем формирования тиоэфирной связи с остатком серосодержащей аминокислоты - цистеином, которая входит в состав ATG-7. Затем он переносится на сопряженный фермент ATG-10, катализирующий образование ковалентной связи с белком ATG-5.
Комплекс ATG-12:ATG-5 вовлекает ATG-16, и вместе белки формируют тримолекулярную структуру. Она также критически важна в аутофагии, поскольку приобретает функцию фермента - лигазы, который задействован во второй убиквитинсопряженной реакции. В этой второй уникальной реакции аргинин, расположенный на С-конце белка ATG-8, убирается белком ATG-4. Теперь уже подготовленный белок ATG-8 активируется ATG-7 для последующего переноса. Наконец две сопряженные системы пересекаются. ATG-12:ATG-5:ATG-16-лигаза способствует присоединению ATG-8 к фосфатидилэтаноламину (к мембране).
Соединенный с липидами ATG-8 является ключевым драйвером удлинения (элонгации) и слияния аутофагосом. Обе сопряженные системы являются высококонсервативными процессами.
У млекопитающих аналог белка ATG-8 называется LC3, и он используется в качестве маркера формирования аутофагосом.
 Есинори Осуми
Есинори Осуми
Японский клеточный биолог, специализация - изучение аутофагии. Работает профессором в Институте инновационных исследований Технологического университета Токио. Образование получил в Университете Токио. В 1974–1977 гг. работал в Университете Рокфеллера, затем вернулся в Японию и получил ставку научного сотрудника. В 1996 г. перешел в Национальный институт фундаментальной биологии (National Institute for Basic Biology), где со временем ему было присуждено звание профессора. С 1998 г. является руководителем исследовательской группы.
Аутофагия и патологические процессы
Изучение молекулярных принципов аутофагии позволило понять патогенез многих заболеваний (рис. 3). В то же время в этой теме еще очень многое остается не изученным.

© Нобелевский комитет по физиологии или медицине (The Nobel Committee for Physiology or Medicine). Иллюстратор: Маттиас Карлен (Mattias Karlén)
Как известно, аутофагия первоначально открыта в виде клеточного ответа на стресс, но сейчас известно, что этот процесс активен и на ранних стадиях дифференциации клеток. В отличие от убиквитин-протеосомной системы, которая в основном деградирует белки с коротким сроком жизни, аутофагия позволяет клетке избавиться от долгоживущих белков. Кроме того, это единственный известный процесс, позволяющий уничтожать целые органеллы, такие как митохондрии, пероксисомы, эндоплазматический ретикулум и прочие. Таким образом, аутофагия играет важнейшую роль в поддержании клеточного гомеостаза. Более того, этот процесс задействован во множестве нормальных процессов, в частности - эмбриогенезе, клеточной дифференциации, которые требуют удаления тех или иных частей цитоплазмы.
Аутофагия прямо или косвенно связана с развитием множества заболеваний.
Онкологические заболевания развиваются несколькими путями и связаны с дифференцировкой клеток. Аутофагия в некоторых случаях защищает организм от развития онкопатологии (онкосупрессор), а в некоторых - способствует развитию злокачественных новообразований. Поскольку аутофагия - это в первую очередь механизм выживания клетки, она помогает выживанию раковых клеток. Это связано с тем, что при изменении фенотипа нормальных клеток, когда они становятся злокачественными, существенно ускоряется скорость их деления и клетки испытывают голодание, гипоксию и пр. В этот момент запускается механизм аутофагии, помогающий клеткам переваривать некоторые участки и выживать в неблагоприятной среде.
Также с помощью аутофагии могут уничтожаться апоптические медиаторы. В таких случаях перспективным терапевтическим направлением представляется ингибирование поздних стадий аутофагии.
Интерес представляет белок Beclin-1 - продукт гена BECN-1 (гомолог дрожжевого гена ATG-6 , регулирующего этапы инициации аутофагии). Экспрессия мутированного белка Beclin-1 характерна для ряда онкологических заболеваний, в частности группы рака молочной железы и яичника. В экспериментах на моделях мышей показано, что гиперэкспрессия нормального Beclin-1 приводит к подавлению опухолевого процесса.
Таким образом, влияние на аутофагию может стать эффективным способом борьбы с онкологическими заболеваниями.
Поскольку интенсивность аутофагии снижается с возрастом, существуют гипотезы, что этот процесс играет ключевую роль в развитии болезней, ассоциированных со старением. По некоторым данным, снижение активности аутофагии связано с развитием остеоартроза.
Одна из наиболее популярных теорий развития нейродегенеративных заболеваний (в настоящее время) также отводит аутофагии значимую роль в этом процессе. Как известно, неправильно упакованные белки имеют тенденцию к формированию нерастворимых агрегатов, являющихся токсичными для клеток. Способность клетки противостоять этому процессу целиком и полностью зависит от активности аутофагии. В исследованиях, проведенных на животных моделях, показано, что активация аутофагии путем ингибирования TOR-киназы приводит к снижению токсичности белковых агрегатов. Кроме того, выключение генов ATG-5 и ATG-7 в нейронах головного мозга мышей приводит к развитию нейродегенеративных заболеваний.
Существует ряд аутосомно-рецессивных нарушений, при которых у человека изменяются процессы аутофагии. Среди них - порок развития мозга, задержки в развитии, умственная отсталость, нарушения двигательных функций, нейродегенерация и другие.
Е. Осуми и его коллеги стали первыми учеными, которые смогли идентифицировать гомологи ATG у млекопитающих. Это позволило проводить исследования аутофагии у высших эукариот.
Со временем были представлены убедительные доказательства того, что аутофагия существует и в клетках всех млекопитающих. Это было подтверждено рядом исследований с использованием электронной микроскопии. Стало известно, что аутофагия присутствует на различных стадиях клеточной дифференциации - от наименее дифференцированных клеток до узкоспециализированных. Тогда было высказано предположение, что аутофагия может быть ответом организма на метаболический стресс, и что процесс может играть роль в патогенезе ряда заболеваний. Со временем было выявлено, что аутофагия существует и в одноклеточных организмах - простейших, что указывало на консервативный характер этого механизма.
Новаторские открытия Е. Осуми существенно повысили интерес к процессу аутофагии. Этот сегмент стал одним из наиболее изучаемых в сфере биомедицинских исследований. Так, в начале 2000-х годов ученые делали не более 150 публикаций в год на эту тему, а в 2015 г. их было более 4 тыс.
На основании фундаментальных открытий Е. Осуми были выделены различные типы аутофагии. Критерием к их разделению послужил объем вещества, который они деградируют. Наиболее изучен процесс макроаутофагии, при котором деградации подвергаются большие объемы цитоплазмы и клеточные органеллы.
Ученые также выяснили, что существует селективная и неселективная аутофагия. Неселективная аутофагия происходит постоянно, в основном она индуцируется стрессом, в частности - голодом. Селективная аутофагия специфических классов субстрата - протеиновых агрегатов, цитоплазматических органелл, вирусов и бактерий, включает специфические адапторы, распознающие субстрат и присоединяющие его к ATG-8/LC3, расположенному на мембране аутофагосомы.
Таким образом, открытие молекулярных основ механизма аутофагии позволило ученым понять очень важный клеточный процесс, характерный для всех эукариот. В настоящее время в этой сфере все еще остается много неизвестного современной науке.
Галина Галковская,
Евгения Бочерикова
© The Nobel Committee for Physiology
or Medicine; фото и иллюстрации
© The Nobel Committee for Physiology or Medicine
Забудьте, наконец, об очищении соком или детоксовых диетах. Нельзя сказать, будто есть что-то плохое в том, чтобы употребить огромное количество измельченной капусты кейл. Однако не ожидайте, что это вымоет токсины из вашего организма быстрее, чем если бы вы питались, как обычно.
Самоканнибализм
Хорошие же новости заключаются в том, что существует малоизвестный способ, позволяющий вам действительно очистить организм, и это тот процесс, который вы можете самостоятельно контролировать. Все, что вам нужно, - это практиковать самоканнибализм. Постойте, что? Нет, вы не ослышались! И да, вы действительно можете обучить свое тело поедать себя. Хотите верьте, хотите нет, но вам это определенно нужно.
Что такое аутофагия?

Естественный процесс, называемый аутофагией (буквально - «самопоедание»), - это встроенная система очистки организма. Ваши клетки создают мембраны, которые охотятся за остатками мертвых, больных или изношенных клеток, пожирают их, разбирают их на запчасти и используют пригодные молекулы для того, чтобы производить энергию.
Очищение тела через переработку ненужных частиц

Вы можете думать об этой системе, как о внутренней программе переработки в вашем организме. Аутофагия делает организм человека более эффективным, так как он сам избавляется от неработающих частиц, останавливает рост раковых клеток, а также препятствует метаболическим дисфункциям, таким как ожирение или диабет.
Польза аутофагии

Существуют также доказательства того, что этот процесс может сыграть важную роль в контроле воспаления и иммунной системы. Когда ученые вывели крыс, у которых отсутствовала способность к аутофагии, они становились толстыми, сонливыми, у них был повышенный уровень холестерина и мозговые нарушения. Если сложить все вместе, то можно смело сказать, что аутофагия - это ключ к замедлению процесса старения, и вы можете научиться тому, как контролировать ее в вашем организме.
Как спровоцировать аутофагию?

Так как же вам стоит поедать самих себя? Это вопрос, который вы вряд ли задавали ранее, но сейчас вы узнаете на него ответ. В первую очередь вам стоит узнать, что аутофагия - это реакция на стресс, так что вам придется довести ваш организм до стрессового состояния, чтобы запустить в более усиленном режиме ваши системы самоканнибализма. В данный момент вы думаете, что эта статья выглядит совсем уж странно, но поверьте: если вы дочитаете ее до конца, то ваша жизнь может серьезно измениться к лучшему. Как обычно и бывает, кратковременный дискомфорт может привести к долговременным преимуществам, поэтому вам стоит ознакомиться с этими тремя наиболее эффективными способами запустить процессы аутофагии.
Упражнения

Вам всегда стоит помнить о том, что упражнения - это первейший способ довести ваш организм до стрессового состояния. Занятия спортом на самом деле наносят урон вашим мышцам, вызывая крошечные микротрещины, которые ваше тело тут же начинает залечивать, что делает ваши мышцы более сильными и более устойчивыми к дальнейшему подобному урону. Занятия спортом - это один из самых популярных способов, которым люди непреднамеренно очищают свой организм. Так что то ощущение свежести и обновленности после тяжелой тренировки - это признак того, что вы очистились. Одно исследование подробно изучило аутофагосомы - структуры, которые формируются вокруг поврежденных клеток, от которых тело решило избавиться. После того как ученые вывели крыс, у которых аутофагосомы светятся ярко-зеленым светом, они обнаружили, что скорость, с которой эти структуры избавляются от поврежденных клеток, у крыс, пробежавших тридцать минут на колесе, значительно возрастает. И скорость продолжала расти даже тогда, когда крысы бежали уже почти полтора часа без перерыва. Но что насчет людей? Определение необходимой нагрузки и продолжительности упражнений, которые запустят процессы аутофагии, а также степени, в которой этот процесс можно регулировать, является сложным вопросом, и на него сейчас не так просто найти точный ответ. Однако можно сказать, что упражнения дают большое количество преимуществ, даже если не учитывать их роль в аутофагии, так что вам в любом случае стоит заниматься спортом. И если вы любите тяжелые упражнения - для вас же лучше. Чем интенсивнее занятия спортом, тем больше выгода.
Голодание

Ирония заключается в том, что у тех, кто любит «очищать» организм с помощью соков и смузи, употребление любой пищи работает против аутофагии. Отказ от еды - это действие, которое принесет вам немало стресса. Вашему организму это определенно не понравится, но в итоге он получит от этого огромную пользу. Более того, ученые уже успели продемонстрировать, что от периодического голодания имеется немало конкретных преимуществ, таких как снижение риска возникновения диабета или сердечно-сосудистых заболеваний, и все они записываются на счет аутофагии.
Также поражает то, как много исследований было сосредоточено на действии этого процесса в головном мозге человека во время голодания. Предполагается, что данный процесс может быть эффективным способом снижения риска возникновения нейродегенеративных заболеваний, таких как болезнь Альцгеймера или Паркинсона. В ходе некоторых исследований было обнаружено, что периодическое голодание улучшает когнитивные функции мозга, структуру мозга, а также нейропластичность, что может позволить мозгу запоминать новую информацию с большей легкостью. Конечно, не было конкретно доказано, что именно аутофагия является этому причиной, а также стоит упомянуть, что испытания проводились на грызунах. Поэтому нельзя сразу же сказать с уверенностью, что у людей будет такая же реакция. Практикующие специалисты говорят, что если вы хотите эффективно голодать, то вам лучше делать это периодически, сессиями от 12 до 36 часов. В процессе голодания рекомендуется ограничивать физическую активность только растяжкой или базовыми занятиями йогой.
Ограничивайте употребление углеводов
Хотя некоторые профессионалы не едят по 18 часов несколько раз в неделю, они признают, что среднестатистическому человеку будет сложно так жить. Простой отказ от еды при любом возможном случае также может оказаться неплохим вариантом - исследования показали, что даже один день голодания в месяц уже снижает риск возникновения сердечно-сосудистых заболеваний. Но существует и другой способ получить все ту же пользу, при этом не отказываясь от вашего любимого стейка (хотя от конфет вам все же придется отказаться). Этот способ называется кетозом - все более популярной диетой среди бодибилдеров и всех тех людей, которые хотя жить дольше. Суть кетоза заключается в том, чтобы снизить содержание в организме углеводов настолько, чтоб у вашего тела не оставалось другого выбора, кроме как использовать жир в качестве источника энергии. Кетоз позволяет людям избавиться от жира, при этом сохраняя мышцы, и существуют доказательства того, что такая диета помогает организму бороться с раковыми опухолями, снижает риск возникновения диабета, а также защищает от возникновения некоторых видов умственных расстройств, в частности от эпилепсии. Исследования показывают, что более половины детей с эпилепсией, которые придерживались этой диеты, начали испытывать приступы в два раза реже. Кетоз - это как обход аутофагии. Вы получаете все те же метаболические изменения и преимущества, что и от голодания, но при этом вам не приходится голодать.
3 октября 2016 года в Стокгольме Нобелевским комитетом был объявлен лауреат премии в номинации по физиологии и медицине. Им оказался Ёсинори Осуми - японский микробиолог, подробно описавший механизм аутофагии клеток живых организмов. Не сказать, что это стало сногсшибательным ноу-хау, ведь данный процесс был открыт ещё в 60-х гг. XX века. Однако талантливому профессору удалось раскрыть его с новой стороны, и одна из самых престижных международных наград оказалась в его руках.
Мало какие научные достижения, за которые ежегодно вручается Нобелевская премия, оказываются достоянием широкой публики. В большинстве своём они интересны лишь узкому кругу знающих и просвещённых. Однако открытие японского учёного с тех пор стало невероятно популярным.
Сегодня о нём говорят не только диетологи, медики и биологи, но и те рядовые граждане, которые ведут здоровый образ жизни, регулярно занимаются чисткой организма, стараются похудеть или являются приверженцами оздоравливающей системы голодания. Оказывается, механизмы клеточной аутофагии в жизни не только людей, но и всех живых существ играют важную роль.
О талантливом учёном
Ёсинори Осуми (родился в 1945 году) - японский учёный, микробиолог, профессор, доктор наук, преподаватель Токийского технологического института, член Европейской Организации Молекулярной Биологии.
Японский учёный, молекулярный биолог Ёсинори Осуми
Начиная с 80-х гг. XX века, занимался изучением исключительно процесса аутофагии. Он был открыт задолго до него, ещё в 60-е гг., но о его роли в организме и важности для человеческой жизни никто даже не подозревал. И только Ёсинори Осуми удалось выяснить в начале 90-х годов, насколько он значим для каждого живого существа на планете.
Задолго до Нобелевской премии японского микробиолога неоднократно награждали за его открытие. Вот некоторые из самых значительных наград:
- 2008 год - премия Асахи, «За точные молекулярные исследования аутофагии и внутриклеточной системы деструкции».
- 2015 - премия Розенстила, «В знак признания за новаторские открытия молекулярных и биологических функций аутофагии».
- 2016 - премия Поля Джанссена, «За открытие молекулярной основы аутофагии как универсального процесса самопереваривания клеток для получения энергии, спасающей жизнь во время голодания».
- 2016 - Нобелевская премия, «За открытие механизмов аутофагии».
- 2017 - премия за прорыв в медицине, «За исследование аутофагии и системы рециркуляции, которую клетки используют для получения питательных веществ из своих собственных несущественных или повреждённых компонентов».
Лишь в одной из формулировок вышеперечисленных премий упоминается . Так почему все считают, что именно за него в 2016 году Ёсинори Осуми дали Нобелевскую премию? Чтобы разобраться в этом вопросе, нужно вникнуть в суть открытия.
Это интересно. В 1974 году Нобелевскую премию получил Кристиан де Дюв - учёный, открывший лизосомы. И он же незадолго до этого ввёл термин «аутофагия».
Аутофагия
Термин восходит к древнегреческому языку и переводится с него как «самопоедание». Он означает процесс ликвидации клетками собственных частиц, которые оказываются вредными, лишними, ненужными. Как уже было сказано, ещё в 60-е годы учёные сделали открытие, что он время от времени запускается, но почему, когда именно, с какой целью и какое это оказывает влияние на работу организма в целом, с тех пор так никто и не выяснил. И лишь спустя 20 лет Ёсинори Осуми взялся за его изучение вплотную.
Материалом для исследования стали дрожжи. Однако всё, что выяснил в ходе своих научных изысканий обладатель Нобелевской премии, относится ко всем живым клеткам, в том числе и человеческого организма. И вот какие открытия он сделал.
В течение жизни под влиянием различных факторов (питания, условий проживания, климата, вредных привычек) в организме накапливается мусор:
- токсины;
- дефектные белки;
- различные вредные вещества;
- омертвевшие частицы;
- инфекции, бактерии, вирусы;
- патологические, повреждённые ткани.
Оказывается, природа наделила клетки способностью самостоятельно избавляться от всего этого. Схематично и упрощённо, не вдаваясь в сложную биологическую терминологию, процесс аутофагии выглядит следующим образом:
Состояние стресса → Клетки распознают в своём составе инородные частицы → Атакуют их → Заключают их в аутофагосомы (похоже на то, как мы складируем мусор в пакеты) → Перемещают их в лизосомы (контейнеры) → Там разрушают и переваривают → Образовавшиеся продукты переработки используют для получения нужной энергии, собственного омоложения и регенерации, внутреннего питания
Результат превосходит все ожидания: мало того, что клетки очищаются от мусора, среди которого оказываются инфекции и вирусы, опасные для здоровья, они ещё и обновляются. Причём никаких внешних ресурсов для этого не требуется.
Как выяснил Ёсинори Осуми, если процессы аутофагии протекают в человеческом организме регулярно и без сбоев, это гарантирует:
- высокую продолжительность жизни;
- замедление процессов старения;
- крепкий иммунитет, отражающий атаки любых, даже самых опасных бактерий и вирусов;
- отличное самочувствие;
- физическую активность и высокие интеллектуальные способности;
- бесперебойную работу всех органов и систем.
А самое главное, что удалось выяснить Нобелевскому лауреату, нарушения в процессе аутофагии влекут за собой такие тяжёлые патологии, как рак, ДЦП, болезни Альцгеймера и Паркинсона, диабет и многие другие, с которыми современная медицина справиться не может. Получается, что, если клетки будут регулярно заниматься «самоедством», риск всех этих заболеваний сводится к нулю.
Собственно, за то, что японский учёный открыл миру истинную причину таких серьёзных патологий (нарушенную аутофагию) и в то же время приоткрыл завесу, как их лечить (наладить этот механизм), он и получил Нобелевскую премию. Остаётся выяснить, а при чём здесь всё-таки голодание?
Аутофагия и голодание
С помощью аутофагии можно продлить жизнь и излечиться от самых тяжёлых заболеваний. Автоматически этого не происходит потому, что запускается она крайне редко. Ёсинори Осуми выяснил, что клетки начинают переваривать свои патогенные части лишь в условиях сильного стресса. И одним из способов его создания является голодание.
Когда клетки получают питание извне (в процессе переваривания съедаемых человеком продуктов), им нет смысла выполнять дополнительную работу - распознавать свои повреждённые элементы и заниматься их утилизацией. Но, как только они в течение длительного времени не получают никакой внешней подпитки, они начинают искать выход из сложившейся ситуации. И единственным спасением становятся те самые лишние частицы, которые можно переработать и потребить, чтобы не погибнуть.
Получается, что, работая над аутофагией, японский учёный неожиданно для своих исследований доказал ещё и пользу голодания. Именно оно запускает этот процесс и, соответственно, гарантирует и долгую жизнь, и избавление чуть ли не от всех заболеваний.
 Процесс клеточной аутофагии
Процесс клеточной аутофагии
Однако стоит сразу оговориться, что в своих исследованиях Ёсинори Осуми подробно описывает исключительно процесс аутофагии: как он запускается, как протекает, что на него влияет, какое значение он имеет для здоровья и жизни человека и иные нюансы. Он не предлагает авторскую систему голодания, как считают многие, подобно , или другим исследователям. Он лишь упомянул о том, что именно воздержание от еды создаёт условия стресса, при котором клетки начинают самоочищаться.
Поэтому нельзя утверждать, что Нобелевскую премию Ёсинори Осуми дали именно за голодание. Нет, он её получил за описание механизмов аутофагии. Но эти два понятия тесно взаимосвязаны, а простому человеку гораздо ближе первое - отсюда и пошло это разночтение.
Значение открытия Ёсинори Осуми
В связи с Нобелевской премией 2016 года в области физиологии и медицины интерес к процессам аутофагии вырос в несколько раз. Практически все современные исследования и труды о голодании ссылаются на открытие Ёсинори Осуми, обосновывая его невероятную пользу. Но здесь нужно уточнить некоторые моменты.
Часто можно услышать, что Ёсинори Осуми получил Нобелевскую премию за однодневное голодание. С одной стороны, мы выяснили, что это косвенно ошибочное заявление. С другой, рациональное зерно в этой формулировке есть, и именно на него должны обратить внимание все те, кто планирует использовать открытие японского учёного в своих целях.
Как его можно использовать?
Кажется, схема максимально проста: запускаем процесс аутофагии с помощью лечебного голодания - и живём без болезней и старости много-много лет. Самые активные уже начали всё это претворять в жизнь: взяли за основу методики 40-дневного воздержания от пищи (системы Джигурды, Суворина) и постятся. Только вряд ли у кого-то получится пройти весь этот путь до конца и добиться желаемых результатов. В чём подвох?
Длительные голодания, предлагаемые большинством существующих методик (Войтовича, Николаева, Лавровой, Щенникова), не поддерживаются официальной медициной. Их польза не только не доказана научно, но и активно опровергается. Риск серьёзных нарушений в работе органов и даже летального исхода слишком высок, чтобы решаться на такое сомнительное оздоровление. Нобелевскую премию за это точно ещё никому не дали.
А вот краткосрочных от 12 часов до 3 дней (не больше!) достаточно для того, чтобы запустить полный цикл процесса аутофагии и добиться желаемых результатов.
К сожалению, пока нет какой-то чёткой концепции или запатентованной методики лечебного голодания, которая бы основывалась конкретно на открытии Ёсинори Осуми. Каков оптимальный срок воздержания, как часто его практиковать, сколько воды можно пить, что разрешено, а что запрещено - все эти вопросы пока остаются открытыми и решаются каждым в индивидуальном порядке.
Те, кто регулярно практикует голодания, советуют брать именно , так как оно и переносится легче, и не вызывает сильного ухудшения самочувствия, и вполне вписывается в рамки выходного дня.
Кому подойдёт?
Аутофагия имеет важное значение для жизни и здоровья каждого человека. Все мы дышим загрязнённым воздухом, едим консерванты и красители и накапливаем в себе настоящие мусорные свалки. Справиться с ними могут и сами клетки, но им в этом нужно помочь. Поэтому абсолютно всем нужно время от времени устраивать краткосрочные голодания.
Но особенно полезными они окажутся для тех, кто:
- страдает от ожирения и лишнего веса (согласно исследованиям, именно жировые клетки накапливают в себе максимальное количество вредных веществ);
- попадает в группу риска по онкологии (из-за наследственного фактора);
- находится в солидном возрасте (после 50 лет это нужно делать регулярно, для профилактики Альцгеймера и Паркинсона);
- планирует в скором времени зачатие малыша (во избежание риска ДЦП).
Несмотря на то, что Ёсинори Осуми получил Нобелевскую премию вовсе не за , как ошибочно считают многие, описанные им механизмы аутофагии тесно с ним связаны. Его открытие даёт надежду абсолютно всем людям на излечение заболеваний, перед которыми до сих пор бессильна современная медицина. Некоторые всерьёз считают, что при правильной реализации этого достижения вполне реально замедлить процессы старения и существенно увеличить продолжительность жизни.
© ПОТАПНЕВ М.П., 2014 УДК 612.014.3.017.1
Потапнев М.П.
АУТОФАГИЯ, АПОПТОЗ, НЕКРОЗ КЛЕТОК И ИММУННОЕ РАСПОЗНАВАНИЕ
своего и чужого
Белорусский государственный медицинский университет Минздрава Республики Беларусь, 220116, Минск
В обзоре литературы представлены данные о роли основных типов клеточной смерти для формирования иммунного ответа на патогены и собственные антигены. Рассмотрены основные механизмы аутофагии, апоптоза и некроза клеток, значение образующихся клеточных продуктов для индукции иммунного ответа. Отмечена роль аутофагии как клеточной автономной системы защиты от патогенов и клеточного стресса. Определена ведущая роль апоптоза, апоптозассоциированных молекулярных образов (паттернов) в индукции иммунологической толерантности. Подчеркнуто решающее значение некроза и продуктов повреждения собственных клеток в индукции воспалительной реакции макроорганизма и эффективного иммунного ответа на собственные антигены, патогены и молекулярные образы (паттерны) патогенов. Обсуждено взаимодействие различных типов клеточной смерти при патологических состояниях.
Ключевые слова: аутофагия; апоптоз; некроз; клеточная смерть; патогены; воспаление; иммунный ответ. Potapnev M.P.
AUTOPHAGY, APOPTOSIS, NECROSIS AND IMMUNE RECOGNITION OF SELF AND NONSELF
Belarusian State Medical University, Ministry of Public Health, 220116, Minsk, Belarus
The review of literature discusses the role of most essential types of cell death (autophagy, apoptosis, necrosis) for induction of immune response to pathogens and self antigens. The main mechanisms of cell death and biological characteristics of cellular products, released during autophagy, apoptosis, necrosis were reported. The role of autophagy as cellular self-defense system against pathogens and cellular stress was underlined. The receptor-ligand interaction for induction of immune tolerance by apoptotic cells and the role of apoptotic cell-associated molecular patterns (ACAMPs) and dendritic cells were described. Brief description of mechanisms of necrotic cell-induced inflammation and immune response as well as leading role of damage-associated molecular patterns/ DAMPs were done. Interaction of DAMPs and pathogen-associated molecular patterns/PAMPs in induction of host defense against pathogens was described. It was concluded that differential type of cell death may be occurred depending on strength of danger signal affecting cells and their function.
Key words: autophagy; apoptosis; necrosis; cell death; pathogens; inflammation; immune response.
Считается, что основной принцип действия иммунной системы заключается в распознавании чужого или измененного своего и его последующем удалении. Классическим примером иммунного распознавания чужого являются реакции врожденного и приобретенного иммунитета против микроорганизмов (бактерий, вирусов). Иммунное распознавание измененного своего ассоциировано с аутоиммунными заболеваниями. С развитием представлений о (за)программированной клеточной смерти (ПКС) стала важной оценка связи иммунитета с поддержанием клеточного гомеостаза в макроорганизме . Всякие изменения клеток в процессе роста и дифференцировки, старения, естественного отмирания, метаболической дисфункции, стресса, воздействия патологического процесса (инфекция, стерильное воспаление) должны рассматриваться иммунной системой как нарушения клеточного гомеостаза. Оценке роли ПКС в запуске иммунных реакций посвящен настоящий обзор.
На основании морфологических и биохимических критериев выделяют три основных типа ПКС: апоптоз (ПКС I типа), аутофагия (ПКС II типа) и некроз (ПКС III типа) . ПКС типов I и II имеют определенные генетически механиз-
Потапнев Михаил Петрович (Potapnev Michael Petrovich), email:[email protected]
мы реализации, поэтому называются активными. ПКС III типа (первичный некроз в результате внешнего повреждения) является неуправляемым, поэтому называется пассивным. Дополнительно выделяют вторичный некроз как конечный результат апоптоза , управляемый некроз (некроптоз) и другие пути гибели клеток . Перечень известных (13) типов клеточной смерти регламентирован Номенклатурным комитетом . Характеристика трех основных типов ПКС представлена в таблице.
Внимание иммунологов к клеточной смерти определяется тем, что не только инфекционные антигены и молекулярные образы (паттерны) патогенов (pathogen-associated molecular patterns - PAMPs), отличающие его от макроорганизма , но и продукты повреждения собственных клеток (damage-associated molecular patterns - DAMPs) вызывают воспаление и иммунный ответ . P. Matzinger подчеркнула, что для иммунной системы важно распознавание и ответ на сигналы опасности, образующиеся в результате повреждения тканей (клеток), а не выяснение различий между своим и чужим.
Аутофагия
Аутофагия - процесс прижизненной утилизации (деградации с помощью лизосом) измененного метаболитами содержимого цитоплазмы для поддержания клеточного и энергетического гомеостаза . Аутофагию рассматривают
ИММУНОЛОГИЯ № 2, 2014
Основные типы клеточной смерти
Характеры- Тип клеточной смерти
стика аутофагия апоптоз некроз
Назначение Деградация и внутриклеточная утилизация поврежденных органелл и белков без вреда для клетки. В случае избыточной деградации -клеточная смерть Деградация отмирающих клеток без воспалительного и иммунного ответа организма Ограничение очага нежизнеспособной ткани путем воспаления и иммунного ответа на токсические и угрожающие организму воздействия
Морфология клеток Вакуолизация цитоплазмы клетки Конденсация и уплотнение клетки, конденсация хроматина, фрагментация ядра, образование апоптоти-ческих телец Набухание органелл с последующим разрывом внутренних и внешних мембран. Набухание и последующий лизис клеток
Механизм действия Последовательное образование в цитоплазме фагофоры, аутофагосомы, аутолизосомы или шаперонопосредованное слияние с лизосомами Каспаззависимый (рецепторный) или митохондриально-зависимый пути деградации ДНК Неконтролируемое повреждение клетки или рецепторзависимый (RAGE, TLRs, CD91 и др.) путь разрушения клетки
Ыаркер LC3-II, ULK 1, ATG12, ATG4, GABARAP ДНК фрагменты 50 kbp, ФС внешней мембраны, FAS, CASP 3, APAF1 ЛДГ, HBGH1, S100-белки, АТФ, HSP90
Участие фагоцитоза Отсутствует Присутствует Присутствует
как преимущественно «запрограммированное выживание клетки» . Стресс вызывает аутофагию, а избыточная активность аутофагии ведет к клеточной смерти . Недостаточность аутофагии провоцирует накопление метаболитов, связанных со старением, дегенеративными процессами в нервной ткани и печени, аутоиммунные, легочные заболевания (особенно на фоне курения). Показана связь аутофагии с болезнью Крона, муковисцидозом, ожирением, сепсисом .
Основной тип аутофагии - макроаутофагия, включающая этапы инициации, нуклеации, элонгации и слияния (с лизосомой) . Измененные белки цитоплазмы (в результате стресса, недостатка энергетического обеспечения), поврежденные митохондрии, избыточный эндоплазматический ретикулум (ЭР), пероксисомы транслоцируются к мембранам органелл благодаря комплексированию с белками ULK 1/2, Atg13, Atg101, fIp-200. На мембранах органелл (ЭР, митохондрии, аппарат Гольджи) эти белки формируют комплекс I, включающий дополнительно белки Vps34, Beclin
I, Vps15, Atg14L. Вокруг комплекса I образуется внутренняя мембрана фагофоры. Формирование аутофагосомы (диаметром 0,3-1 мкм) с двойной мембраной требует участия LC3
II, образующегося в результате липолизации фосфатидилэ-таноламином цитозольного белка LC3, и комплекса белков Atg5-Atg12/Atg16L1. Последующее созревание аутофагосомы в аутофаголизосому осуществляется путем слияния с лизосомами с помощью комплекса белков II, включающего Vps34, Beclin 1, UVRAG . В аутофаголизосоме осуществляется деградация измененных белков под действием гидролаз и высвобождение в цитоплазму питательных и энергоемких субстанций . Кроме макроаутофагии выделяют микроаутофагию (когда захват содержимого цитоплазмы осуществляется путем инвагинации мембраны лизо-сом) и шаперонопосредованную аутофагию (когда доставка цитоплазматического материала в лизосомы осуществляется с помощью белков-шаперонов) .
В связи с наличием в цитоплазме клетки измененных своих и чужеродных макромолекул процесс аутофагии, являясь метаболическим, выступает еще как механизм распознавания и утилизации внутриклеточных микроорганизмов (вирусы, бактерии, простейшие), несущих PAMPs . Проникновение в цитоплазму микроорганизмов и их продуктов запускает механизмы аутофагии в качестве клеточной автономной защитной системы-cell-autonomous defense system. Разделение цитоплазмы клетки на отдельные, ограниченные (эндо)мембранами участки и органеллы (т. е. компартментализация) предполагает наличие в каждом их них своего набора рецепторов, распознающих чужеродные PAMPs и измененные собственные DAMPs. Это создает многоступенчатую систему защиты от патогенов, проник-
ших внутрь клетки. На каждом этапе продвижения патогена в клетке происходит распознавание ДНК, агрегированных собственных белков, комплекса микробов и сывороточных белков. Патоген сталкивается с различными ферментами; NO и H2O2; наличием или недостатком питательных веществ . Микробы активируют рецепторы на эндомембранах цитоплазмы, что ведет к формированию инфламмасомы, продукции интерлейкина (ИЛ)-1р и ИЛ-18 . Попадание патогена в аутофаголизосомы резко изменяет условия его существования за счет действия рН, гидролаз, супероксидных анионов. При этом возможны персистенция патогена (длительная для M. tuberculosis, короткая для других бактерий) в аутофагосомах либо разрушение патогена в аутофаголизо-сомах . Toll-like receptors (TLRs) распознают попавшие в цитоплазму макрофагов бактериальный липополисахарид (ЛПС), вирусную однонитчатую рибонуклеиновую кислоту (онРНК), другие полимерные нуклеиновые кислоты. При аутофагии в распознавании внутриклеточных патогенов (Str. pyogenes, M. tuberculosis, BCG, Salmonella, вирусы) участвуют TLRs, RLRs (retinoid acid inducible gene I-like receptors), NLRs (nucleotide oligomerization domain- like receptors) . TLR3, распознающий РНК вирусов, локализуется в эндосомах клетки; TLR7, TLR8, TLR9, распознающие РНК и ДНК вирусов и бактерий, CpG-мотивы нуклеиновых кислот микробного происхождения, - в эндолизосомах. RLRs, распознающие РНК вирусов, и NLRs, распознающие PAMPs (мурамил дипептид, токсины, кристаллы солей, другие компоненты) бактерий, вирусов, клеточные продукты химического воздействия и УФ-облучения, расположены в цитоплазме . Важной функций TLRs является обеспечение жесткого контроля за нормальной (комменсальной) микрофлорой кишечника .
PAMPs, распознаваемые TLR1, TLR2, TLR4, TLR5, TLR6, вызывают образование в инфламасоме цитокинов воспаления ИЛ-ф и ИЛ-18. PAMPs, распознаваемые TLR7, TLR9, стимулируют продукцию интерферона-а (ИФНа) и ИФНр, что способствует формированию Th1 иммунного ответа . Продукция ИЛ-1Р и ИЛ-18 защищает клетки от вируса гриппа и бактерий рода Shigella соответственно. А вызванный в результате активации инфламмасом пироптоз (гибель клеток с признаками апоптоза и некроза) губителен для сальмонелл, легионелл и других бактерий . Активация TLR4 разрушает связь Bcl-2 c белком Beclin 1, что ведет к образованию фагосомы из фагофоры . Активация TLRs индуцирует быстрый переход Lc3 из цитоплазмы в фагосому, активацию клетки, способствует созреванию фагосомы и слиянию ее с лизосомой . L. monocytogenesis в цитоплазме клетки распознают NLRs и TLR2, а S. flexneri распознают NLRs, что приводит к деградации микробов механизмами аутофагии с участием инфламмасом . При захвате
живых бактерий (в отличие от мертвых) в инфицированную клетку попадает микробная мРНК, которая создает дополнительный сигнал опасности (vita-PAMPs), активирующий инфламмасомы типа NLRP3 и TRIF-зависимую продукцию ИФНр . Таким образом, аутофагия выступает как механизм деградации микроорганизмов при их попадании в цитоплазму клетки и распознавании патогенассоциированными рецепторами.
Аутофагия участвует в презентации антигенов Т-клеткам. Образование протеосом, ассоциированных с ЭР, или ау-тофагосом создает благоприятные условия для контакта мембранно-связанных молекул MHC I или II классов с пептидами и последующей передачи их комплексов на внешнюю мембрану антигенпредставляющих клеток для индукции соответственно CD8- или CD4- зависимых Т-клеточных реакций . Белки аутофагии LC3 и GABARAP в аутофагосомах повышают в 20 раз сродство собственных и чужеродных пептидов к молекулам MHC II класса . Блокирование гена аутофагии Atg5 подавляет образование CD4+ Т-клеточного (Th1) ответа на вирус простого герпеса или ВИЧ-1, а также препятствует распознаванию В-клеток, инфицированных вирусом Эпштейна-Барр .
Аутофагия в тимическом эпителии является основой негативной селекции аутореактивных Т-клеток. Блок гена аутофагии Atg5 приводит к аутоиммунному CD4+ Т-клеточному пролиферативному заболеванию мышей и накоплению апоптотических CD4+ и CD8+ Т-клеток . Дефицит аутофагии в периферических Т-клетках вызывает ускоренную клеточную смерть наивных, но не Т-клеток памяти, что связывают с продукцией супероксидных анионов при активации наивных Т-клеток . Важной функцией аутофагии является изоляция поврежденных митохондрий, генерирующих супероксидные анионы, как источник стресса и повреждения (вплоть до гибели) самой клетки .
Аутоиммунный ответ при сахарном диабете и аутоиммунном гепатите вызывают аутоантигены GAD65 (глутамат декарбоксилаза 65) и SMA (мутантная к-легкая цепь иммуноглобулинов), которые подвергаются в цитоплазме шаперо-нопосредованной аутофагии с участием HSC70 и связанного с лизосомами мембранного белка LAMP-2A соответственно. После деградации в лизосомах они вместе с молекулам MHC II класса презентируются аутореактивным cD4+ Т-клеткам. Образование в аутофаголизосомах цитрулированных пептидов под действием пептидиларгинин деаминаз и формирование их комплексов с молекулами MHc II класса является основой аутоиммунного cD4+ Т-клеточного ответа при ревматоидном артрите - РА . В Т-клетках мышей линии MRL с лимфопролиферативным синдромом, аналогом системной красной волчанки (СКВ) человека, выявляется значительное количество аутофагосом в Т-клетках, что объясняют их длительным выживанием .
Продукция супероксидных анионов митохондриями макрофагов способствует переваривание бактерий в процессе аутофагии . Бактерии, распознаваемые NLRs, стимулируют аутофагию в фибробластах. В дендритных клетках (ДК) это приводит к представлению пептидов бактерий вместе с молекулами MHC II класса CD4+ T-клеткам . Важной защитной функцией аутофагии является способность снижать уровень собственных DAMPs в цитоплазме и сдерживать секрецию ИЛ-ф и ИЛ-18 в ответ на экзогенные источники DAMPs. Механизмы аутофагии обеспечивают деградацию инфламмасом - комплекса белков, превращающих прокаспазу-1 в каспазу-1, конвертирующую про- ИЛ-ф и про-ИЛ-18 в секретируемые активные цитокины . Блокировка гена аутофагии Atg16L1 приводит у мышей к повышенной продукции ИЛ-ф и ИЛ-18, воспалению, повышению уровня смертности при антигенной стимуляции декстран сульфатом .
Внеклеточные цитокины влияют на процессы аутофагии бактерий и их переваривание в фаголизосомах. Цитокины ТЫ-зависимого ответа ИФНу и фактор некроза опухолей а (ФНОа) стимулируют аутофагию. Цитокины №2-зависимого
ответа ИЛ-4 и ИЛ-13, наоборот, снижают образование фаго-лизосом и повышают внутриклеточное выживание M. tuberculosis . Дифференцировка Т-клеток в Th1 и Th2 in vitro характеризуется большим и меньшим образованием ауто-фагосом соответственно. Внутриклеточные инфекционные агенты (цитомегаловирус, ВИЧ, вирус герпеса простого I, вирус гриппа А, йерсинии, листерии, шигеллы, сальмонеллы, E. coli и др.) избегают иммунного ответа путем ослабления процесса аутофагии .
Аутофагия является физиологическим процессом самообновления клетки, которое при стрессовых воздействиях может привести к ее гибели . В то же время естественное отмирание клеток (у человека от 50 до 500 млрд клеток ежедневно) осуществляется преимущественно путем апоптоза .
Апоптоз. Апоптоз обеспечивает удаление отмирающих клеток посредством фагоцитоза без воспаления, губительного для макроорганизма, или сопровождает очаг воспаления для его ограничения и окончательного заживления . Формирование иммунной системы и созревание антигенспецифических Т- и В-лимфоцитов также сопровождается массовым апоптозом клеток . Апоптоз обеспечивает поддержание клеточного гомеостаза, стимуляцию клеточной регенерации, заживление ран. Апоптотические клетки (АК) утилизируются соседними клетками эпителия, эндотелия, фибробластами, макрофагами, ДК . При заболеваниях и переливании хранившейся донорской крови в периферической крови, лимфоузлах, костном мозге выявляются апоптотические тельца диаметром 0,2 мкм, образующиеся из АК. Выделяемые АК липидные медиаторы (лизофосфа-тидилхолин, сфингозин-1-фосфат), рибосомальный dRP S19, EMAP II эндотелиальных клеток, TyrRS синтетазу, тромбоспондин 1, растворимый рецептор к ИЛ-6, фракталкин (CX3-CR1L), нуклеотиды АТФ и УТФ привлекают фагоциты. При этом лактоферрин, выделяемый клетками слизистых и нейтрофилов при апоптозе, избирательно подавляет хемотаксис нейтрофилов, но не макрофагов . Поверхностная экспрессия фосфатидилсерина (ФС), других окисленных липидов и калретикулина является признаком ранних АК, распознаваемых рецепторами макрофагов (стабилин-2, CR3, рецепторы-мусорщики (scavenger receptors), CD91, CD31, TIM4, CD36, steroid receptor activator 1; TAM-рецепторы (Ty-ro2, Ax1, Mer); LRP-1). Молекулярные маркеры АК получили общее название apoptotic cell-associated molecular patterns (ACAMPs) . Макрофаги распознают апоптотические клетки посредством нескольких апоптозассоциированных рецепторов одновременно для быстрого удаления клеток на ранних этапах апоптоза. Экспрессия поверхностного CD31 (и/или CD47) на АК предотвращает их захват макрофагами . Важно, что рецепторы макрофагов, распознающие АК и апоптотические тельца, отличаются от рецепторов, распознающих PAMPs и DAMPs . Более того, активация рецепторов, различающих АК и апоптотические тельца, способствует подавлению распознавания макрофагами PAM-Ps инфекционных агентов через TLRs .
Распознавание АК и апоптотических телец облегчается участием сывороточных опсонинов Gas6, MFG-E8, P2GP1, аннексина I, С-реактивного белка (СРБ), пентраксина PTX-3, коллектинов, dq-компонента комплемента, сурфактантов SP-A и SP-D (в легочной ткани) и т. д. . При этом опсо-нин MFG-E8, участвующий в захвате АК макрофагами, одновременно подавляет фагоцитоз некротических клеток (НК) и их иммуногенность для ДК. C1q взаимодействует с ФС ранних АК, а коллектин маннозасвязывающий лектин (MBL) - с поздними АК. Калретикулин (в комплексе с CD91), пентрак-сины СРБ, SAP (компонент сывороточного амилоида Р); фи-колины взаимодействуют с поздними АК . Оценивая роль системы комплемента и естественных антител в клиренсе АК. Ряд авторов определили, что лизофосфатидилхо-лин, появляющийся (и частично секретируемый) на поверхности АК, является мишенью естественных антител - IgM, а также маннозасвязывающих белков, других коллектинов. Их взаимодействие в свою очередь приводит к связыванию
ИММУНОЛОГИЯ № 2, 2014
с C1q, C3b/bi. В результате АК фагоцитируются без активации выброса макрофагами провоспалительных цитокинов. Аутоиммунные реакции с участием антикардиолипиновых антител класса G, наоборот, протекают с участием комплемента и аутоантител к фосфолипидам мембран поздних АК . Важно, что апоптотические тельца на ранних этапах апоптоза покрыты элементами ФС-содержащей внешней мембраны клеток, а на поздних этапах - элементами эндоплазматических мембран. И если антигенная презентация ранних апоптотических телец вызывает образование иммунорегуляторных Т-клеток (Treg), то контакт поздних апопто-тических телец с ДК вызывает образование ТЫ7-клеток . Апоптотические нейтрофилы (и внешние мембраны лизированных нейтрофилов) вызывают продукцию трансформирующего ростового фактора в (ТРФр) макрофагами, а внутреннее содержимое лизированных нейтрофилов - образование ИЛ-8, ФНОа, хемокина MIP-2 . В очаге воспаления сами нейтрофилы проявляют «каннибализм», фагоцитируя апоптотические нейтрофилы (например, индуцированные УФ-облучением). Этому способствуют дополнительная активация TLRs эффекторных нейтрофилов и цитокины ФНОа и гранулоцитарно-макрофагальный колониестимулирующий фактор (ГМ-КСФ), но не ИЛ-1-р, ИЛ-6, ИЛ-8, ИЛ-12, ИЛ-17 . В очаге воспаления макрофаги являются основными фагоцитами АК . Это не приводит к продукции провоспалительных цитокинов (ИЛ-1р, ФНОа, ИЛ-6, ИЛ-12), но вызывает образование иммуносупрессорных ИЛ-10, ТРФр, простагландина Е2 (ПГЕ2) . Формируется иммунная толерантность к антигенам АК и одновременно к другим антигенам, включая PAMPs микроорганизмов, которая опосредуется СЭ8а+ДК. ДК, стимулированные АК, представляют антиген(ы) только CD8+ Т-клеткам, а ДК, стимулированные НК, представляют антиген(ы) CD4+ и CD8+ Т-клеткам . Иммуносупрессия, развивающаяся в результате массового образования АК и их захвата макрофагами, лежит в основе лечебного действия экстракорпорального фотофереза у пациентов с хроническими воспалительными заболеваниями .
Длительно протекающий процесс апоптоза в очаге воспаления может привести к формированию фиброза, что связывают со способностью макрофагов, фагоцитировавших АК, секретировать ТРФр и другие ростовые факторы . В то же время подавление воспаления, усиление репаративных процессов при фагоцитозе АК приводит при наличии генетической предрасположенности к аутоиммунным заболеваниям (СКВ, хроническое обструктивное заболевание легких) . В норме В1-подобные клетки с фенотипом CD43+CD27-IgM+ или cD24++cD38++cD27- IgM+ являются основным источником естественных антител к поверхностным молекулам АК . Значительное количество АК в герминативных центрах лимфоузлов у пациентов с СКВ обеспечивает длительное выживание и костимуляцию аутореактивных В-клеток, активированных однониточной ДНК, нуклеосомами, другими клеточными антигенами. Это связано с Oq-зависимым генетическим дефектом быстрого клиренса ранних АК и накоплением поздних АК с признаками вторичного некроза . Образующиеся низкоаффинные антитела класса IgM взаимодействуют с клетками, находящимися на ранних стадиях апоптоза, а высокоаффинные антитела класса IgG - с клетками, находящимися на поздних стадиях апоптоза. Плазмацитоидные ДК и активация ДНК-связывающих TLR9 В-клеток обеспечивает Т-независимое образовании аутоантител. Индуцируемая АК продукция иммуносупрессорного ИЛ-10 значительно снижена при стимуляции В-клеток иммунными комплексами, включающими хроматин, или апоптотическими тельцами, образующимися на поздних этапах апоптоза .
Элиминация АК осуществляется в основном на ранних этапах апоптоза, когда экспрессия на внешней мембране ФС и калретикулина сигнализирует об «измененном своем» . Ранние этапы апоптоза обратимы, их продление обеспечивает фагоцитоз большинства АК и формирование толерантности иммунной системы . Переход клеток на поздние этапы
апоптоза характеризуется снижением уровня гликозилирова-ния поверхностных молекул, фрагментаций ядерной ДНК и признаками вторичного некроза, вызывающего воспаление и иммунный ответ .
Основными путями запуска апоптоза клеток являются рецепторный (extrinsic), обусловленный внешним воздействием, или стрессиндуцированный (intrinsic), связанный с внутренним воздействием. Рецепторный путь запуска апоптоза клетки опосредован рецепторами смерти (death receptors), включающими Fas, TNFR (рецептор I типа к ФНОа), TRAIL, Apo2/Apo3. Активация каспаз является ключевой для апоптоза и последовательность их включения достаточно описана в литературе . Стрессиндуцированный (митохондриальный) путь апоптоза связан с высвобождением цитохрома С из митохондрий и регулируется белками семейства Bcl2. Каспаззависимая активация и повышение уровня супероксидных анионов (преимущественно за счет повреждения митохондрий) определяют иммуносупрессорное действие АК. Толерогенное действие АК, считается, опосредуется Heg-клетками, вызывающими TRAIL-индуцированную гибель CD4+ Т-клеток-хелперов [ 52]. Оба пути апоптоза приводят к поверхностной экспрессии ФС, фрагментации ДНК ядра, образованию апоптотических телец и их быстрому фагоцитозу. Это предотвращает иммунный ответ на отмирающую клетку, продукцию макрофагами цитокинов воспаления, презентацию ДК клеточных антигенов.
При инфицировании клетки проявляют признаки раннего апоптоза (экспрессия на клеточных мембранах ФС, начало фрагментации ДНК) и NF-кБ-зависимого пути клеточной активации. Одновременно клетки сдерживают репликацию патогенов без образования DAMPs, свойственных некротическим клеткам . Дефекты звеньев апоптоза (преимущественно митохондриальнозависимого пути активации), или запоздалый запуск апоптоза приводят к распространению инфекции (вызванной Legionella pneumonia, Pseudomonas aeroginosa, Helicobacter pylori), сепсису . Многие вирусы содержат ингибиторы каспаз, а Chlamydiae и Coxiella burnetii блокируют выход цитохрома c из митохондрий и апоптоз клетки, что обеспечивает жизненный цикл патогена на ранних этапах инфекции . Захват АК, содержащих бактерии, вызывает созревание ДК, воспаление, полноценный (Th17) иммунный ответ, при захвате неинфицированных АК признаки созревания ДК и воспаления отсутствуют, формируется иммуносупрессия . Стратегия ограниченной репликации патогена в АК выгодна отсутствием сильного иммунного ответа на некроз клеток и массовый выброс бактерий во внеклеточное пространство .
Некроз. Клетки, отмирающие в результате травмы, дегенеративных процессов, воздействия патогена, эффективно утилизируются путем некроза. Некроз демаркирует нежизнеспособную ткань, подлежащую уничтожению и последующему восстановлению . Некроз клеток всегда сопровождается воспалением и ведет к выраженному иммунному ответу и последующей репарации тканей . НК характеризуются разрушением внешней клеточной мембраны и поступлением во внеклеточное пространство скрытых внутриклеточных молекул (см. таблицу), что вызывает токсическую реакцию окружающих здоровых клеток и иммунный ответ . Первичный некроз клеток не зависит от действия каспаз и является прямым результатом внешнего травматического повреждения или запрограммированных генетически событий, связанных с повреждением белка митохондриального матрикса циклофилина Д; воздействием на рецепторы смерти или TLR3/TLR4 и рецепторнезависимым повреждением ДНК . Оксидативный стресс клетки, активные формы кислорода являются индукторами (управляемого) некроза . Вторичный некроз - это конечный результат позднего апоптоза, часто он лежит в основе аутоиммунной патологии (СКВ и другие).
НК фагоцитируются путем макропиноцитоза после исчезновения поверхностных CD31- и СЭ47-молекул, блокирующих фагоцитоз. НК в отличие от АК вызывают созревание ДК
и (Th1) иммунный ответ . НК выделяют внутриклеточные молекулы, провоцирующие воспаление и иммунную реакцию, поэтому они названы аларминами (alarmins) или DAMPs . Они привлекают нейтрофилы в очаг некроза. НК выделяют белки теплового шока (HSP70, HSP90, gp96), калгра-нулины, цитокины (ИЛ-1а, ИЛ-6), формилпептиды митохондрий, РНК, двунитчатую (геномную) ДНК, другие молекулы . Выделение ядерного белка HMGB1 (high-mobility group box 1), связанного в норме с хроматином, является основным маркером (первичного) некроза клеток . При апоптозе и вторичном некрозе HMGB1 удерживается в ядре или находится в цитоплазме или внеклеточно в неактивном (окисленном) состоянии в результате действия супероксидных анионов . Сам HMGB1 является митогеном и хемоаттрактантом, но образуемые им комплексы с однонитчатой ДНК, ЛПС бактерий, нуклеосомой вызывают секрецию макрофагами цитокинов воспаления ФНОа, ИЛ-1р, ИЛ-6, хемокины ИЛ-8, MIP-1a, MIP-ip . Высокий уровень HMGB1 в крови связан с массивным некрозом клеток организма и является маркером системного воспаления . HMGB1 - мощный адъювант образования высокоаффинных антител и созревания ДК . Циркулирующий в кровотоке неокисленный (активный) HMGB1 взаимодействует c TLR2, TLR4, TLR9 и RAGE (receptor for advanced glycation end-products) фагоцитов, вызывая воспалительный ответ . Одновременно HMGB1 (а также HSPs) взаимодействует с CD24 и Siglec-10 на поверхности фагоцитов, что ограничивает воспаление, вызванное DAMPs, но не PAMPs . Разграничение иммунного ответа на патогенассоциированные PAMPs и связанные с повреждением собственных клеток DAMPs происходит на уровне рецепторов клеток. Типичным рецептором для DAMPs является RAGE на клетках иммунной и нервной систем, эндотелиальных клетках, кардиомиоцитах. RAGE распознает белки и липиды, модифицированные в результате неферментативного гликозили-рования и появляющиеся при хронических воспалительных заболеваниях как результат окислительного стресса. RAGE распознает такие продукты НК, как HMGB1 и калгранулины (белки семейства S 100) .
НК выделяют нуклеиновые кислоты. При этом РНК становится двунитчатой, взаимодействует с TLR3 на ДК, а двунитчатая ДНК - с TLR9 фагоцитов, что приводит к продукции ИФНу, CXCL10 (IP-10), ИЛ-1Р, экспрессии костимулирую-щих молекул (cD40, cD54, cD69, MHc II класса) на поверхности макрофагов и ДК . Для того чтобы не вызвать воспаления, молекулы ДНК подвергаются ферментативному расщеплению, например каспаз при апоптозе . Дефект ДНКаз, разрезающих двунитчатую ДНК, вызывает у мышей аутоиммунные заболевания (СКВ, полиартрит). Нуклеотиды АТФ и УТФ, в норме находящиеся в цитоплазме, при некрозе клеток выделяются во внеклеточное пространство. Действуя на пуринэргические рецепторы ДК, они вызывают хемотаксис незрелых ДК, образование NALP3 инфламмасом и секрецию ИЛ-1р, Th2 иммунного ответа . Действие АТФ на аллергенактивированные миелоидные ДК провоцирует развитие легочной аллергии и поддержание бронхиальной астмы . Ядерные рибонуклеопротеины (их короткие фрагменты) выделяются при разрушении НК и выступают в качестве DAMPs, стимулируя образование цитокинов и а-хемокинов . Соли ураты, образуемые из мочевой кислоты при разрушении в цитоплазме эндогенной ядерной или микробной ДНК и ионов натрия внеклеточного пространства, стимулируют образование инфламмасом в макрофагах и ДК, синтез цитокинов ИЛ-1Р, ИЛ-18, ИЛ-33, нейтрофильную инфильтрацию, созревание ДК, усиление антигенспецифического Т-клеточного ответа .
Стрессиндуцированные цитоплазматические белки-шапероны HSP70, HSP90 при некрозе (но не апоптозе) клеток поступают во межклеточное пространство. Внеклеточные HSP70, HSP90 стимулируют образование цитокинов воспаления (ФНОа, ИЛ-1Р, ИЛ-6, ИЛ-12) . Антигенспецифический иммунный ответ на комплекс пептид-HSP значительно возрастает. Клеточными рецепторами HSPs выступают cD91,
CD40, TLR2/TLR4/CD14, рецепторы-мусорщики, LOX-1 . НК выделяют калгранулины (S100 белки), которые распознаются RAGE рецепторами клеток эндотелия, микроглии, моноцитов и становятся маркерами воспаления (при пневмониях, полиартритах и т. д.) . Выделение цитокинов (ИЛ-1, ИЛ-6, ИЛ-33) также может быть результатом стрессового воздействия на клетки и их некротической гибели . Высвобождаемые из НК протеазы и биологически активные молекулы воздействуют на окружающие ткани и отщепляют от них низкомолекулярные фрагменты (гиалуроновая кислота, фибриллярный белок, коллаген, гепарансульфат), которые также вызывают воспаление .
Как и при утилизации АК, сывороточные факторы (кол-лектин MBL) связываются с НК, усиливая их распознавание и связывание с калретикулином на поверхности макрофагов . Макрофаги распознают некротические клетки посредством TLRs, лектин С-типа рецепторов Clec9A, RAGE; CD14, CD91, CD40, Mincle (взаимодействующим с SAP-130) и других. Важно, что рецепторы фагоцитов, распознающие НК, не распознают АК и (частично) распознают молекулы (PAMPs) патогенов (микобактерии, грибки и др.) .
Регулируемый некроз (некроптоз) клеток связан с активностью киназ RIPK1 и RIPK3, проявляется быстрым повышением проницаемости клеточных мембран и высвобождением во внеклеточное пространство внутриклеточных DAMPs . Некроптоз клеток кожи, слизистых, лейкоцитов при ишемической реперфузии вызывает сильный воспалительный ответ. Одновременно он выступает в качестве защитного механизма при вирусной инфекции (при наличии вирусных ингибиторов каспазы 8), а также участвует в поддержании гомеостаза Т-лимфоцитов . Некроптоз инфицированной клетки означает резкое изменение среды обитания внутриклеточных патогенов, что губительно для них. Пироптоз клеток, имея черты апоптоза и некроза, характеризуется образованием инфламмасом как комплекса активированных каспаз и продуцентов цитокинов воспаления ИЛ-1Р и ИЛ-18. Пироптоз эффективно защищает клетки от S. aureus, S. ty-phimurium, P. aeruginosa, L. pneumophila, F.tularensis, B. an-thracis. При этом разные типы специализированных инфлам-масом образуются в ответ на живые бактерии, их токсины, ЛПС, споры, флагеллин, ДНК, РНК вирусов и бактерий . Некроз клеток характеризует продвинутые (не ранние) этапы инфекционного процесса, когда патогены (Shigella, Salmonella, Yersinia, M.tuberculosis) переходят от тактики выживания в апоптотических клетках к тактике разрушения клетки и межклеточного распространения .
Вторичный некроз как исход апоптоза клеток характеризуется выделением DAMPs нуклеосом (фрагментов геномной ДНК размером 180 пар оснований), HMGB1. Иммуностиму-
Индукция «сигналами опасности» различных типов гибели клеток. Сплошные линии - основное действие, пунктирная линия - дополнительное действие (при слабом воздействии), -I означает подавление клеточной гибели. Остальные обозначения в тексте.
ИММУНОЛОГИЯ № 2, 2014
лирующее действие таких DAMPs связано с образованием комплексов нуклеосом с HMGB1, характерных для пациентов с СКВ . Вторичный некроз сопровождается массовым высвобождением модицифированных (в результате ферментативной обработки, окисления) аутоантигенов, которые в комплексе с HSPs (и другими DAMPs) вызывают антигенспецифический иммунный ответ. Но только наличие генетической предрасположенности приводит к формированию аутоиммунной патологии .
Взаимодействие между путями клеточной смерти.
Аутофагия и апоптоз клеток рассматривают как механизмы поддержания жизнеспособности многоклеточного организма, а формирование инфламмасом и некрозиндуцированное воспаление считают механизмами ограниченного отмирания тканей для сохранения макроорганизма . Распознавание DAMPs при аутофагии создает дополнительную страховку клеткам макроорганизма в защите от патогенов с неизвестными PAMPs . В результате инфицирования макрофагов L. pneumophila активация инфламмасом вызывает пироптоз и ау-тофагию, которая защищает клетку от пироптоза и патогена. Но недостаточность аутофагии для противодействия патогену ведет инфицированную клетку к пироптозу . Запуск PIRK1-3-зависимого механизма некроптоза предполагает первоначально высокий уровень аутофагии поврежденных митохондрий и при ее неэффективности последующую деградацию клетки . Аутофагия выступает в качестве механизма утилизации фагоцитированных апоптотические телец макрофагами и ДК. Повышение при некрозе клеток уровня HMGBT цитоплазме стимулирует вместе с HSP27 аутофагию (митофагию) митохондрий и подавляет апоптоз. Другие DAMPs (АТФ, белки S100/ калгранулины, двунитчатая ДНК), взаимодействуя с TLRs, также стимулируют аутофагию в очагах апоптоза. Известно, что основной Beclin 1 -зависимый путь аутофагии (макроаутофагия) может быть подавлен анти-апоптотическими белками семейства Bcl-2 и образованием NLRP3-инфламмасом, т. е. возрастание резистентности клетки к апоптотической гибели повышает ее устойчивость к избыточной аутофагии, приводящей к гибели клеток . При фагоцитозе клеток, умерших путем аутофагии или апоптоза, воспаление отсутствует. Блокирование аутофагии в клетке приводит к накоплению в цитоплазме поврежденных митохондрий, супероксидных анионов, активации NALP3-инфламмасомы, воспалению. Взаимодействие DAMPs с рецепторами RAGE стимулирует аутофагию и подавляет апоптоз клеток . При недостаточном выделении DAMPs из НК в очаге повреждения апоптотические клетки вызывают состояние толерантности и снижение воспаления. ^зревание ДК вызывают DAMPs из НК, но не ACAMPs из АК. Макрофаги, фагоцитировавшие АК, выделяют ТРФр, что вызывает образование Teg-клеток. При фагоцитозе АК, инфицированных E. coli, макрофаги выделяют ТРФр и ИЛ-6, что ведет к образованию ТЫ7-клеток, а при фагоцитозе НК -Th1 иммунного ответа . При совместном воздействии PAMPs и DAMPs последние выполняют роль адъюванта . Известно, что в зависимости от дозы воздействия (например, ФНОа) клетка погибает путем апоптоза (при низких концентрациях) или некроза (при высоких концентрациях) . Связь между апоптозом и некрозом клеток определяется также наличием промежуточных подтипов клеточной смерти - не-кроптоза и других .
Разные типы клеточной смерти как результат ответа клеток на внешние (включая микроорганизмы) и внутренние воздействия могут протекать одновременно и регулировать друг друга (см. схему). До конца неясны механизмы, определяющие выбор пути клеточной смерти, но чем сильнее воздействие, тем сильнее ответ в виде некроза клеток, мощной воспалительной и иммунной реакции макроорганизма. Слабые воздействия (за счет аутологичных apoptotic cell-associated molecular patterns (AcAMPs) или DAMPs, PAMPs нормальной микрофлоры) вызывают интенсификацию аутофагии и апоптоза клеток без очевидной воспалительной и иммунной реакций.
Заключение. Гибель клеток макроорганизма (человек,
животные), обусловленная внешними или внутренними причинами, вызывает иммунный ответ на повреждение. При этом микробные воздействия всегда дозированы концентрацией и жизнеспособностью патогена, его растворимыми продуктами, локализацией очага повреждения . Сочетанное действие PAMPs и DAMPs, наиболее часто встречающееся в реальных условиях, а также влияние толерогенных апопто-тических клеток на их взаимодействие требуют дальнейшего изучения и оценки иммунологических последствий .
литература
1. Ярилин А.А. Апоптоз. Природа феномена и его роль в целостности организма. Патологическая физиология. 1998; 2: 38-48.
3. Бра М., Квинан Б., Сузин С.А. Митохондрии в программированной гибели клетки: различные механизмы гибели. Биохимия. 2005; 70 (2): 284-93.
4. Черников В.П., Белоусова Т.А., Кактурский Л.В. Морфологические и биохимические критерии клеточные гибели. Архив патологии. 2010; 72 (3): 48-54.
5. Galluzzi L., Vitale I., Abrams J.M., Alnemri E.S., Baehrecke E.H., Blagosklonny M.V et al. Molecular definition of cellular death subroutines: recommendations of the Nomenclature Committee on Cell Death 2012. Cell Death Different. 2012; 19 (1): 107-20.
9. Манских В.Н. Пути гибели клетки и их биологическое значение. Цитология. 2007; 49 (11): 909-15.
11. Хаитов Р.М., Пащенков М.В., Пинегин Б.В. Роль паттернораспознающих рецепторов во врожденном и адаптивном иммунитете. Иммунология. 2009; 1: 66-76.
15. Romao S., Gannage M., Munz C. Checking the garbage bin for problems in the house, or how autophagy assists in antigen presentation to the immune system. Semin. Cancer Biol. 2013; 23 (5): 391-6.
16. Rubinsztein D.C., Marino G., Kroemer G. Autophagy and aging. Cell. 2011; 146 (5): 682-95.
19. Walsh C.M., Edinger A.L. The complex interplay between au-tophagy, apoptosis and necrotic signals promotes T-cell homeostasis. Immunol. Rev. 2010; 236 (1): 95-109.
20. Amre D.K., Mack D.R., Morgan K., Krupoves A., Costea I., Lam-brette P. et al. Autophagy gene ATG16L1 but not IRGM is associated with Crohn’s disease in Canadian children. Inflamm. BowelDis. 2009; 15 (4): 501-7.
21. Salminen A., Kaarniranta K., Kauppinen A. Beclin 1 interactome controls the crosstalk apoptosis, autophagy and inflammasome activation: impact on the aging process. Ageing Res. Rev 2012; 12 (2): 520-34.
24. Mostowy S., Cossart P. Bacterial autophagy: restriction or promotion of bacterial replication? Trends Cell Biol. 2012; 22 (6): 283-91.
25. Randow F., MacMicking J.D., James L.C. Cellular self-defense:
how cell-autonomous immunity protects against pathogens. Science. 2013; 340 (6133): 701-6.
26. Lamkanfi M., Dixit v.M. Manipulation of host cell death pathways during microbial infections. Cell Host Microbe. 2010; 8 (l): 44-54.
30. Бонаренко В.М., Лиходед В.Г. Распознавание комменсальной микрофлоры образраспознающими рецепторами в физиологии и патологии человека. Журнал микробиологии, эпидемиологии и иммунологии. 2012; 3: 82-9.
31. Paul-Clark M.J., George P.M., Gatheral T., Parzych K., Wright W.R., Crawford D. et al. Pharmacology and therapeutic potential of pattern recognition receptors. Pharmacol. Ther 2012; 135 (2): 200-15.
40. Byrne B.G., Dubuisson J.-F., Joshi A.D., Persson J.J., Swanson M.S. Inflammasome components coordinate autophage and pyroptosis as macrophage response to infection. mBio.2013; 4 (1): e00620-
12. Available at http://mbio.asm.org/content/4/1/e00620-12.full. pdf+html
41. Kleinnijenhuis J., Oosting M., Platinga T.S., van der Meer J.W.M., Joosten L.A.B., Crevel R.V et al. Autophagy modulates the Mycobacterium tuberculosis-induced cytokine response. Immunology. 2011; 134 (3): 341-8.
42. Гариб Ф.Ю., Ризопулу А.П. Взаимодействие патогенных бактерий с врожденными иммунными реакциями хозяина. Инфекция и иммунитет. 2012; 2 (3): 581-96.
47. Saas P., Angelot F., Bardiaux L., Seilles E., Garnache-Ottou F., Per-ruche S. Phosphatidylserine-expressing cell by-products in transfusion: a pro-inflammatory or an anti-inflammatory effects? Transfus. Clin. Biol. 2012; 19 (3): 90-7.
54. Miles K., Heaney J., Sibinska Z., Salter D., Savill J., Gray D. et al. A tolerogenic role for Toll-like receptor 9 is revealed by B-cell interaction with DNA complexes expressed on apoptotic cells. Proc. Natl Acad. Sci. USA. 2012; 109 (3): 887-92.
59. Проскуряков С.Я., Габай В.Л., Коноплянников А.Г. Некроз -управляемая форма программируемой клеточной смерти. Биохимия. 2002; 67 (4): 467-91.
63. Blander J.M., Sander L.E. Beyond pattern recognition: five immune checkpoints for scaling the microbial threat. Nature Rev. Immunol. 2012; 12 (3): 215-25.
1. Yarilin A.A. Apoptosis. Nature of the phenomenon and its role in the whole organism. Patologicheskaya fiziologiya. 1998; 2: 38-48 (in Russian).
2. Green D.R. The end and after: how dying cells impact the living organism. Immunity. 2011; 35 (4): 441-5.
3. Bras M., Queenan B., Susin S.A. Programmed cell death via mitochondria: Different modes of dying. Biokhimiya. 2005; 70 (2): 231-9 (in Russian).
4. Chernikov V.P., Belousova T.A., Kaktursky L.V. Morphological and biochemical criteria for cell death. Arkhiv patologii. 2010; 72 (3): 48-54 (in Russian).
5. Galluzzi L., Vitale I., Abrams J.M., Alnemri E.S., Baehrecke E.H., Blagosklonny M.V. et al. Molecular definition of cellular death subroutines: recommendations of the Nomenclature Committee on Cell Death 2012. Cell Death Different. 2012; 19 (1): 107-20.
6. Peter C., Wesselborg S., Herrman M., Lauber K. Dangerous attraction: phagocyte recruitment and danger signals of apoptotic and necrotic cells. Apoptosis. 2010; 15 (9): 1007-28.
7. Kaczmarek A., Vandenabeele P., Krysko D.V. Necroptosis: the release of damage-associated molecular patterns and its physiological relevance. Immunity. 2013; 38 (2): 209-23.
8. Rock K.L., Lai J.-J., Kono H. Innate and adaptive immune responses to cell death. Immunol. Rev. 2011; 243 (1): 191-205.
9. Manskikh V.N. Pathways of cell death and their biological importance. Tsitologiya. 2007; 49 (11): 909-15 (in Russian).
10. Janeway C.A. Jr., Medzhitov R. Innate immune recognition. Ann. Rev. Immunol. 2002; 20 (1): 197-216.
11. Khaitov R.M., Pashchenkov M.V., Pinegin B.V. The role of pattern-recognizing receptors in congenital and active immunity. Immunologiya. 2009; 1: 66-76 (in Russian).
12. Seong S.Y., Matzinger P. Hydrophobicity: an ancient damage-associated molecular pattern that initiates innate immune responses. Nature Rev. Immunol. 2004; 4 (6): 469-78.
13. Chen G.Y., Nunez G. Sterile inflammation: sensing and reacting to damage. Nature Rev. Immunol. 2010; 10 (12): 826-37.
14. Kuballa P., Nolte W.M., Castoreno A.B., Xavier R.J. Autophagy and the immune system. Ann. Rev. Immunol. 2012; 30: 611-46.
15. Romao S., Gannage M., Munz C. Checking the garbage bin for problems in the house, or how autophagy assists in antigen
ИММУНОЛОГИЯ № 2, 2014
presentation to the immune system. Semin. Cancer Biol. 2013; 23 (5): 391-6.
16. Rubinsztein D.c., Marino G., Kroemer G. Autophagy and aging. Cell. 2011; 146 (5): 682-95.
17. Tang D., Kang R., Coyne C.B., Zeh H.J., Lotze M.T. PAMPs and DAMPS: signal Os that spur autophagy and immunity. Immunol. Rev. 2012; 249 (1): 158-75.
18. Zelenay S., Reis e Sousa C. Adaptive immunity after cell death. Trends Immunol. 2013; 34 (7): 329-35.
19. Walsh C.M., Edinger A.L. The complex interplay between autophagy, apoptosis and necrotic signals promotes T-cell homeostasis. Immunol. Rev. 2010; 236 (1): 95-109.
20. Amre D.K., Mack D.R., Morgan K., Krupoves A., Costea I., Lambrette P. et al. Autophagy gene ATG16L1 but not IRGM is associated with Crohn’s disease in Canadian children. Inflamm. Bowel Dis. 2009; 15 (4): 501-7.
21. Salminen A., Kaarniranta K., Kauppinen A. Beclin 1 interactome controls the crosstalk apoptosis, autophagy and inflammasome activation: impact on the aging process. Ageing Res. Rev. 2012; 12 (2): 520-34.
22. Levine B., Mizushima N., Virgin H.W. Autophagy in immunity and inflammation. Nature. 2011; 469 (7330): 323- 35.
23. Liu G., Bi Y., Wang R., Wang X. Self-eating and self-defense: autophagy controls innate immunity and adaptive immunity. J. Leukoc. Biol. 2013; 93 (4): 511-9.
24. Mostowy S., Cossart P. Bacterial autophagy: restriction or promotion of bacterial replication ? Trends Cell Biol. 2012; 22 (6): 283-91.
25. Randow F., MacMicking J.D., James L.C. Cellular self-defense: how cell-autonomous immunity protects against pathogens. Science. 2013; 340 (6133): 701-6.
26. Lamkanfi M., Dixit V.M. Manipulation of host cell death pathways during microbial infections. Cell Host Microbe. 2010; 8 (1): 44-54.
27. Mintern J.D., Villadangos J.A. Autophagy and mechanisms of effective immunity. Front. Immunol. 2012; 3: 60.
28. Travassos L.H., Carneiro L.A.M, Ramjeet M., Hussey S., Kim Y.-G., Magalhaes J.G. et al. Nod1 and Nod2 direct autophagy by recruiting ATG16L1 to the plasma membrane at the site of bacterial entry. Nature Immunol. 2010; 11 (1): 55-62.
29. Kumar H., Kawai T., Akira S. Pathogen recognition by the innate immune system. Int. Rev. Immunol. 2011; 30 (1): 16-34.
30. Bondarenko V.M., Likhoded V.G. Recognition of commensal microflora by pattern recognition receptors in human physiology and pathology. Zhurnal Mikrobiologii, epidemiologii i immunologii. 2012; 3: 82-9 (in Russian).
31. Paul-Clark M.J., George P.M., Gatheral T., Parzych K., Wright W.R., Crawford D. et al. Pharmacology and therapeutic potential of pattern recognition receptors. Pharmacol. Ther. 2012; 135 (2): 200-15.
32. Strowig T., Henao-Mejia J., Elinav E., Flavell R. Inflammasomes in health and disease. Nature. 2012; 481 (7381): 278-86.
33. Underhill D.M., Goodridge H.S. Information processing during phagocytosis. Nature Rev. Immunol. 2012; 12 (7): 492-502.
34. Sander L.E., Davis M.J., Boekschoten M.V., Amsen D., Dascher C.C., Ryffel B. et al. Detection of prokaryotic mRNA signifies microbial viability and promotes immunity. Nature. 2011; 474 (7351): 385-9.
35. Schmid D., Pypaert M., Munz C. Antigen- loading compartments for major histocompatibility complex class II molecules continuously receive input from autophagosomes. Immunity. 2007; 26 (1): 79-92.
36. Paludan C., Schmid D., Landthaler M., Vockerodt M., Kube D., Tuschl T. et al. Endogenous MHC class II processing of a viral nuclear antigen after autophagy. Science. 2005; 307 (5709): 593-6.
37. Pua H.H., Guo J., Komatsu M., He Y.W. Autophagy is essential for mitochondrial clearance in mature T lymphocytes. J. Immunol. 2009; 182 (7): 4046-55.
38. Lu J.V., Walsh C.M. Programmed necrosis and autophagy in immune function. Immunol. Rev. 2012; 249 (1) : 205-17.
39. Gros F., Arnold J., Page N., Decossas M., Korganow A.-S., Martin T. et al. Macroautophagy is deregulated in murine and human lupus T lymphocytes. Autophagy. 2012; 8 (7): 1113-23.
40. Byrne B.G., Dubuisson J.-F., Joshi A.D., Persson J.J., Swanson M.S. Inflammasome components coordinate autophage and pyroptosis as
macrophage response to infection. mBio. 2013; 4 (1): e00620-12. Available at http://mbio.asm.org/content/4/1/e00620-12.full.pdf+html
41. Kleinnijenhuis J., Oosting M., Platinga T.S. , van der Meer J.W.M., Joosten L.A.B., Crevel R.V et al. Autophagy modulates the Mycobacterium tuberculosis-induced cytokine response. Immunology. 2011; 134 (3): 341-8.
42. Garib F.Yu., Rizopulu A.P. Interaction of pathogenic bacteria with innate immune reactions of host. Infektsiya i immunitet. 2012; 2 (3): 581-96 (in Russian).
43. Majai G., Petrovski G., Fesus L. Inflammation and the apopto-phagocytic system. Immunol. Lett. 2006; 104 (1-2): 94-101.
44. Janssen W.J., Henson P.M. Cellular regulation of the inflammatory response. Toxicol. Pathol. 2012; 40 (2): 166-73.
45. Zitvogel L., Kepp O., Kroemer G. Decoding cell death signals in inflammation and immunity.Cell. 2010; 140 (6): 798-804.
46. Bekeredjian-Ding I. B cell encounters with apoptotic cells. Autoimmunity. 2013; 46 (5): 307-11.
47. Saas P., Angelot F., Bardiaux L., Seilles E., Garnache-Ottou F., Perruche S. Phosphatidylserine-expressing cell by-products in transfusion: a pro-inflammatory or an anti-inflammatory effects? Transfus. Clin. Biol. 2012; 19 (3): 90-7.
48. Jeannin P., Jaillon S., Delneste Y. Pattern recognition receptors in the immune response against dying cells. Curr. Opin. Immunol. 2008; 20 (5): 530-7.
49. Lauber K., Blumenthal S.B., Waibel M., Wesselborg S. Clearance of apoptotic cells: getting rid of the corpses. Mol. Cell. 2004; 14 (3): 277-87.
50. Fadok V.A., Bratton D.L., Guthrie L., Henson P.M. Differential effects of apoptotic versus lysed cells on macrophage production of cytokines: role of proteases. J. Immunol. 2001; 166 (11): 6847-54.
51. Hellberg L., Fuchs S., Gericke C., Sarkar A., Behhen M., Solbach W. et al. Proinflammatory stimuli enhance phagocytosis of apoptotic cells by neutrophil granulocytes. Scient. World J. 2011; 11: 2230-6.
52. Ferguson T.A., Choi J., Green D.R. Armed response: how dying cells influence T-cell functions. Immunol. Rev. 2011; 241 (1): 77-88.
53. Douglas I. S., Diaz del Valle F., Winn R.A., Voelkel N.F. P-catenin in the fibroproliferative response to acute lung injury. Am. J. Respir. Cell Mol. Biol. 2006; 34 (3): 274-85.
54. Miles K., Heaney J., Sibinska Z., Salter D., Savill J., Gray D. et al. A tolerogenic role for Toll-like receptor 9 is revealed by B-cell interaction with DNA complexes expressed on apoptotic cells. Proc. Natl Acad Sci. USA. 2012; 109 (3): 887-92.
55. Ashida H., Mimuro H., Ogawa M., Kobayashi T., Sanada T., Kim M. et al. Cell death and infection: a double-edged sword for host and pathogen survival. J. Cell Biol. 2011; 195 (6): 931-42.
56. Manfredi A.A., Capobianco A., Bianchi M.E., Rovere- Querini P. Regulation of dendritic- and T-cell fate by injury-associated endogenous signals. Crit. Rev. Immunol. 2009; 29 (1): 69-86.
57. Torchinsky M.B., Garaude J., Martin A.P., Blander J.M. Innate immune recognition of infected apoptotic cells directs T(H)17 cell differentiation. Nature. 2009; 458 (7234): 78-82.
58. Bianchi M.E. HMGB1 loves company. J. Leukoc. Biol. 2009; 86 (3): 573-6.
59. Proskuryakov S.Ya., Gabai V.L., Konoplyannikov A.G. Necrosis - an active, regulated form of programmed cell death (review). Biokhimiya. 2002; 67 (4): 467-91 (in Russian).
60. Idzko M., Hammad H., van Nimwegen M., Kool M., Willart M.A.M., Muskens F. et al. Extracellular ATP triggers and maintains asthmatic airway inflammation by activating dendritric cells. Nature Med. 2007; 13 (8): 913-9.
61. Kono H., Rock K.L. How dying cells alert the immune system to danger. Nature Rev. Immunol. 2008; 8 (4): 279-89.
62. Eigenbrod T., Park J.-H., Harder J., Iwakura Y., Nunez G. Cutting edge: Critical role for mesothelial cells in necrosis-induced inflammation through the recognition of IL-1a released from dying cells. J. Immunol. 2008; 181 (2): 8194-8.